
11. Эволюция живого


Эволюция органического мира Земли неразрывно связана с эволюцией литосферы. История развития литосферы Земли подразделяется на геологические эры: катархейскую, архейскую, протерозойскую, палеозойскую, мезозойскую, кайнозойскую. Каждая эра делится на периоды и эпохи. Геологическим эрам, периодам и эпохам соответствуют определенные этапы развития жизни на Земле.
Катархей, архей и протерозой объединяются в криптозой - «эпоху скрытой жизни». Ископаемые остатки криптозоя представлены отдельными фрагментами, не всегда поддающимися идентификации. Палеозой, мезозой и кайнозой объединяются в фанерозой - «эпоху явной жизни». Начало фанерозоя характеризуется появлением скелетообразующих животных, хорошо сохраняющихся в ископаемом состоянии: раковинных моллюсков, древних членистоногих.
|
Эон |
Эра |
Период |
Начало, млн лет назад |
Продолжительность, млн лет |
|
Фанерозой |
Кайнозой |
Антропогеновый |
2-по н. в. |
2 |
|
Неогеновый |
25-2 |
23 |
||
|
Палеогеновый |
70-25 |
45 |
||
|
Мезозой |
Меловой |
135-65 |
70 |
|
|
Юрский |
195-135 |
60 |
||
|
Триасовый |
230-195 |
35 |
||
|
Палеозой |
Пермский |
280-230 |
50 |
|
|
Каменноугольный |
350-280 |
70 |
||
|
Девонский |
410-350 |
60 |
||
|
Силурийский |
440-410 |
30 |
||
|
Ордовикский |
500-440 |
60 |
||
|
Кембрийский |
570-500 |
70 |
||
|
Криптозой |
Протерозой |
Эдиакаранский |
680-570 |
110 |
|
1500-700 |
800 |
|||
|
2500-1500 |
1000 |
|||
|
Архей |
Архейский |
3000-2500 |
500 |
|
|
Катархей |
Катархейский |
4500-3000 |
1500 |
Ранние этапы развития органического мира
Предшественники современных организмов (архебионты) характеризовались наличием основных компонентов клетки. Существовали системы обмена веществ (электрон-транспортные цепи) и системы воспроизведения, передачи и реализации наследственной информации (репликация нуклеиновых кислот и биосинтез белка на основании генетического кода).
Дальнейшее развитие органического мира включает эволюцию отдельных групп организмов в составе экосистем. Таким образом, на ранних этапах развития органического мира должны были сформироваться основные способы питания: фотоавтотрофный, гетеротрофный голозойный и гетеротрофный сапротрофный. Фотоавтотрофный тип питания включает поглощение неорганических веществ поверхностью тела и последующий хемосинтез или фотосинтез. При гетеротрофном сапротрофном типе питания происходит поглощение растворенных органических веществ всей поверхностью тела, а при гетеротрофном голозойном типе питания - захват крупных пищевых частиц и их переваривание.
В условиях избытка готовых органических веществ гетеротрофный сапротрофный способ питания является первичным. Большая часть архебионтов специализировалась именно на гетеротрофном сапротрофном питании. У них формируются сложные ферментные системы. Это привело к увеличению объема генетической информации, появлению ядерной оболочки, разнообразных внутриклеточных мембран и органоидов движения. У части гетеротрофов происходит переход от сапротрофного питания к голозойному.
Другая часть архебионтов специализировалась на автотрофном питании. Древнейшим способом автотрофного питания является хемосинтез. На основе ферментно-транспортных систем хемосинтеза возникает фотосинтез. Избыток углеводов, образующихся при фиксации СО2, позволил синтезировать разнообразные полисахариды.
Вероятно, на ранних стадиях эволюции органического мира Земли был широко распространен обмен генами между совершенно разными организмами. В ходе синтезогенеза свойства гетеротрофных и фотоавтотрофных организмов объединились в одной клетке. Это привело к формированию различных отделов водорослей.
11.2. Основные этапы эволюции растений
Водоросли - многочисленная неоднородная группа первично-водных фотоавтотрофных организмов. В ископаемом состоянии водоросли известны еще из докембрия (свыше 570 млн лет назад), а в протерозое и начале мезозоя уже существовали все ныне известные отделы.
В конце силура (≈ 400 млн лет назад) возникают высшие (наземные) растения.
В силуре происходило обмеление океана и опреснение воды. Содержание кислорода в атмосфере до появления наземных растений было значительно ниже современного. При дефиците кислорода лимитирующим фактором в атмосфере является ультрафиолет. Выход растений на сушу сопровождался развитием метаболизма фенольных соединений (дубильных веществ, флавоноидов, антоцианов), которые участвуют в осуществлении защитных реакций, в том числе от мутагенных факторов (ультрафиолет, ионизирующие излучения, некоторые химические вещества).
Продвижение растений на сушу связано с появлением ряда ароморфозов (ароморфоз - прогрессивное эволюционное изменение строения, приводящее к общему повышению уровня организации и интенсификации функций живых организмов):
-
появлением дифференцированных тканей: покровных, проводящих, механических, фотосинтезирующих;
-
появлением дифференцированных органов: побега (органа углеродного питания) и корня (органа минерального питания);
-
существенными изменениями в обмене веществ.
Дальнейшая эволюция высших растений разделилась на две линии: гаметофитную и спорофитную.
Представители гаметофитной линии - современные моховидные. Это бессосудистые растения, у которых отсутствуют специализированные проводящие и механические ткани.
Другая линия эволюции привела к появлению сосудистых растений, у которых имеются все ткани высших растений. Благодаря появлению всех типов тканей происходит дифференцировка тела растений на корень и побег. В течение девона формируются современные группы споровых растений (плауны, хвощи, папоротники).
В начале мезозоя (≈ 220 млн лет назад) появляются первые голосеменные растения. В кайнозое голосеменные уступают господство покрытосеменным. В настоящее время покрытосеменные представлены множеством жизненных форм: деревьями, кустарниками, лианами, однолетними и многолетними травами, водными растениями. Особого разнообразия достигает строение цветка, что способствует точности опыления и обеспечивает интенсивное видообразование - к покрытосеменным относится около 250 тыс. видов растений.

11.3. Основные этапы эволюции животных
Эукариотические организмы, специализирующиеся на гетеротрофном питании, дали начало животным и грибам. Первые животные были представлены одноклеточными организмами. Многие из них занимали промежуточное положение между животными, водорослями и грибами. Упрощенная схема эволюции животных приведена на рис. 48.
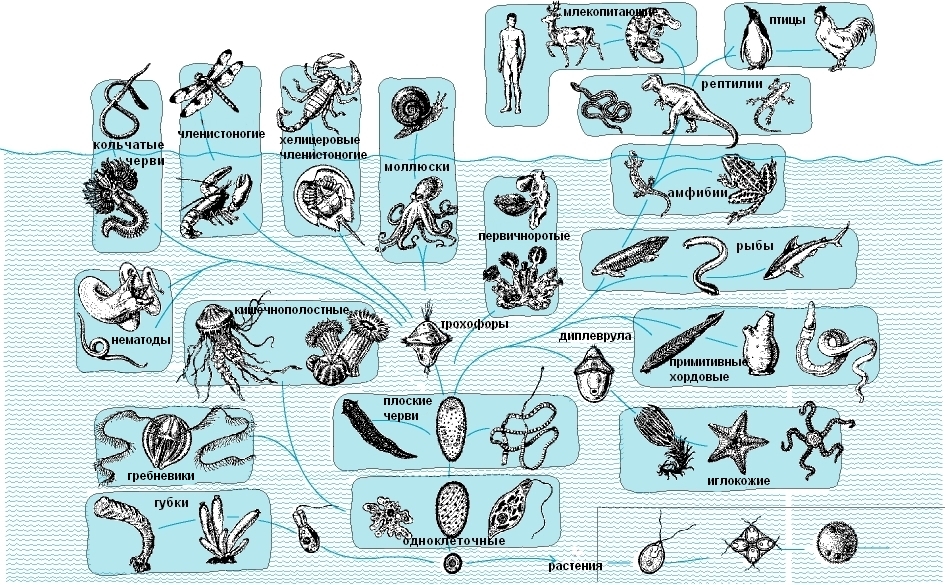 |
|
Рис. 48. Упрощенная схема эволюции животных |
В протерозойской эре возникают все известные типы многоклеточных беспозвоночных животных. С появлением цветковых растений в меловом периоде начинается совместная эволюция насекомых и цветковых (коэволюция), и у них формируются совместные адаптации (коадаптации). В кайнозойской эре насекомые, как и цветковые растения, находятся в состоянии биологического прогресса.
От примитивных хордовых животных в силуре происходят первые позвоночные (бесчелюстные). Низшие челюстноротые позвоночные представлены разнообразными рыбами. Современные классы рыб (хрящевые и костные) формируются в конце палеозоя - начале мезозоя).
Часть костных рыб благодаря легочному дыханию и появлению настоящих конечностей - дала начало первым четвероногим - амфибиям (земноводным). Первые земноводные вышли на сушу в девонском периоде, но их расцвет приходится на каменноугольный период. Современные амфибии появляются в конце юрского периода.
Параллельно среди четвероногих появляются организмы с зародышевыми оболочками. Благодаря зародышевым оболочкам, а также ряду других признаков рептилии полностью утратили зависимость от воды. Появление первых примитивных рептилий относится к концу каменноугольного периода. В перми появляются разнообразные группы рептилий: зверозубые, первоящеры и другие. В начале мезозоя формируются ветви черепах, плезиозавров, ихтиозавров. Начинается расцвет рептилий.
От групп, близких к первоящерам, отделяются две ветви эволюционного развития. Одна ветвь в начале мезозоя дала начало многочисленной группе псевдозухий. Псевдозухии дали начало нескольким группам: крокодилам, птерозаврам, предкам птиц и динозаврам. Вторая ветвь в начале мелового периода привела к появлению подкласса чешуйчатых (ящерицы, хамелеоны и змеи).
Однако рептилии не смогли утратить зависимость от низких температур: теплокровность у них невозможна из-за неполного разделения кругов кровообращения. В конце мезозоя с изменением климата происходит массовое вымирание рептилий.
Лишь у части псевдозухий в юрском периоде появляется полная перегородка между желудочками, редуцируется левая дуга аорты, происходит полное разделение кругов кровообращения, и становится возможной теплокровность. В дальнейшем эти животные приобрели ряд адаптаций к полету и дали начало классу птицы. В юрских отложениях мезозойской эры (≈ 150 млн лет назад) обнаружены отпечатки первоптиц: археоптерикса и археорниса. Вероятно, это были древесно-лазающие животные, которые могли планировать, но не были способны к активному полету. Современные группы птиц появляются только в начале кайнозойской эры.
Расцвет птиц в кайнозойской эре связан с рядом крупных идиоадаптаций (появление перьевого покрова, специализация опорно-двигательного аппарата, развитие нервной системы, забота о потомстве и способность к перелетам), а также с рядом признаков частичной дегенерации (например, утрата зубов).
В начале мезозойской эры появляются первые млекопитающие. В юрском периоде мезозойской эры млекопитающие были представлены, как минимум, пятью классами. Один из этих классов, вероятно, дал начало современным первозверям, а другой - сумчатым и плацентарным. Плацентарные млекопитающие благодаря появлению плаценты и настоящего живорождения в кайнозойской эре переходят в состояние биологического прогресса.
Исходным отрядом плацентарных являются насекомоядные. От насекомоядных рано отделились неполнозубые, грызуны, приматы и ныне вымершая группа креодонтов - примитивных хищников. От креодонтов отделились две ветви. Одна из этих ветвей дала начало современным хищным, от которых отделились ластоногие и китообразные. Другая ветвь дала начало примитивным копытным, а затем непарнокопытным, парнокопытным и родственным отрядам.
Окончательная дифференцировка современных групп млекопитающих завершилась в эпоху великих оледенений - в плейстоцене.
В конце кайнозойской эры у части приматов возникает особый тип ароморфоза - переразвитие коры больших полушарий головного мозга. В результате возникает совершенно новый вид организмов - человек разумный.
11.4. Происхождение человека и общества
Где, когда и как появился наш прямой предок, человек разумный - Homo sapiens? Где и когда появились мы, современные люди, - Homo sapiens sapiens? Точных ответов на эти вопросы мы, скорее всего, никогда не получим, но споры на эту тему ведутся давно. Постоянно делаются новые открытия, появляются новые находки все более древних или более похожих на нас форм. Ведутся генетические исследования. Не проходит и месяца, чтобы в каком-нибудь научном журнале не появилась статья, утверждающая, что наконец-то получены данные, позволяющие решить проблему происхождения нашего вида.
Долгое время научные дискуссии о происхождении человека были уделом археологов и палеонтологов. На основании находок ископаемых остатков и их изучения был сделан вывод о том, что род Homo возник в Африке примерно 2,5 млн лет назад, а около 1,8 млн лет назад его представители распространились и в Европе, и в Азии. Так что к моменту появления человека разумного все части мира, за исключением Австралии и Америки, уже были населены представителями нашего рода.
Остатки же современного человека, найденные на сегодняшний день в Азии (за исключением Ближнего Востока), имеют возраст приблизительно 60 тыс. лет. В Европе современные люди появились максимум 40 тыс. лет назад, а в Америке - еще позже. Какое отношение имели древние люди, жившие в разных частях света, к более позднему появлению в тех же местах человека разумного? Можно ли называть их нашими предками и если да, то в какой степени? Возникшая в 30-х годах. XX века полицентрическая гипотеза (ее еще называют мультирегиональной) постулирует, что популяции представителей рода Homo, обитавшие на разных континентах, в частности в Африке и Азии, эволюционировали в сторону нынешних Homo sapiens sapiens независимо. То есть современные люди появились независимо друг от друга во многих местах, причем процесс расхождения их предков от общего ствола начался 500-700 тыс. лет назад, а может быть, и раньше.
 Однако
с точки зрения теории Дарвина вероятность того, что два разных вида,
эволюционируя в одно и то же время и в двух удаленных друг от друга местах,
придут к одному «результату», крайне низка. Принимая мультирегиональную
гипотезу, мы должны признать, что предки современных людей происходили, по
крайней мере, от очень ограниченного числа видов Homo, а скорее всего от
одного. То есть, несмотря на начавшуюся давно независимую эволюцию и изначальные
морфологические отличия, все мы происходим все-таки от одного вида.
Однако
с точки зрения теории Дарвина вероятность того, что два разных вида,
эволюционируя в одно и то же время и в двух удаленных друг от друга местах,
придут к одному «результату», крайне низка. Принимая мультирегиональную
гипотезу, мы должны признать, что предки современных людей происходили, по
крайней мере, от очень ограниченного числа видов Homo, а скорее всего от
одного. То есть, несмотря на начавшуюся давно независимую эволюцию и изначальные
морфологические отличия, все мы происходим все-таки от одного вида.
В противоположность полицентрической гипотезе происхождения человека была сформулирована моноцентрическая гипотеза, известная также под названием «Из Африки». Согласно моноцентрической гипотезе, род Homo на протяжении своей эволюционной истории был представлен многими независимо эволюционировавшими видами и формами. Но только одна из этих групп, возникшая, как считается, в Африке, дала начало современным людям. Остальные, включая тех, кто успел широко расселиться по земному шару, вымерли, не оставив следа.
В последнее время с появлением и развитием методов молекулярно-генетического анализа, в дискуссии палеонтологов и археологов включились генетики. Надо заметить, что из самих ископаемых остатков, к сожалению, пока не удается выделить ДНК, пригодную для исследования - речь идет об анализе распределения различных генов в популяциях современного человека и попытках реконструировать ситуации, при которых такое распределение могло сформироваться в ходе истории. При этом выводы генетиков далеко не всегда отвечают предположениям, сделанным на основе морфологических исследований ископаемых костей. Так, генетический анализ показывает, что время расхождения азиатских и африканских популяций человека не может быть столь большим, как это предполагали палеонтологи-полицентристы.
Третья гипотеза, названная «сетевой», или гипотезой «гибридизации-замещения», была предложена в 1990-е годы Г. Брауером из Гамбургского университета. Не отвергая возможность африканского происхождения современного человека, эта гипотеза предполагает участие в становлении нашего вида и генов других представителей рода Homo - за счет скрещиваний, происходивших во время миграций и расселения.
 Впервые
связь человека с приматами показал К. Линней: он рассматривал человека как
биологический вид Homo sapiens. Ч. Дарвин в своей работе «Происхождение
человека и половой отбор» показал, что наши анатомические, физиологические,
онтогенетические и поведенческие характеристики весьма сходны с таковыми у
приматов. Дарвин установил, что происхождение человека как биологического вида
подчиняется тем же закономерностям, что и происхождение других биологических
видов.
Впервые
связь человека с приматами показал К. Линней: он рассматривал человека как
биологический вид Homo sapiens. Ч. Дарвин в своей работе «Происхождение
человека и половой отбор» показал, что наши анатомические, физиологические,
онтогенетические и поведенческие характеристики весьма сходны с таковыми у
приматов. Дарвин установил, что происхождение человека как биологического вида
подчиняется тем же закономерностям, что и происхождение других биологических
видов.
Особенности эволюции человека заключаются в том, что параллельно с образованием нового биологического вида шло формирование человеческого общества. Поэтому происхождение человека и общества представляет собой единый процесс - антропосоциогенез.
Становление человека как социального существа не могло осуществиться без определенных биологических предпосылок, которые существовали уже у отдаленных предков человека - древних приматов:
-
приматы - относительно крупные организмы и, как следствие, имеют крупный головной мозг (сотни кубических сантиметров);
-
в связи с древесно-лазающим образом жизни у них имеется бинокулярное цветовое зрение и хватательные конечности;
-
приматы - стадные животные, что создало предпосылки для формирования общества.
Отряд приматы отделился от примитивных насекомоядных в конце мелового периода мезозойской эры (свыше 70 млн лет назад). Полуобезьяны и обезьяны разделились около 50-60 млн лет назад. Широконосые обезьяны (Южная Америка) и узконосые обезьяны (Африка и Азия) разделились около 35-40 млн лет назад.
От общего ствола узконосых обезьян примерно 23 млн лет назад отделились человекообразные обезьяны (гоминоиды). Примерно 12 млн лет назад человекообразные обезьяны разделились на азиатских (современные гиббон и орангутан) и африканских (современные горилла и шимпанзе, у которых объем мозга не превышает 460 см3).
Примерно 7-9 млн лет назад (на границе плиоцена и миоцена) от африканских гоминоидов отделяется ветвь гоминид (людей).
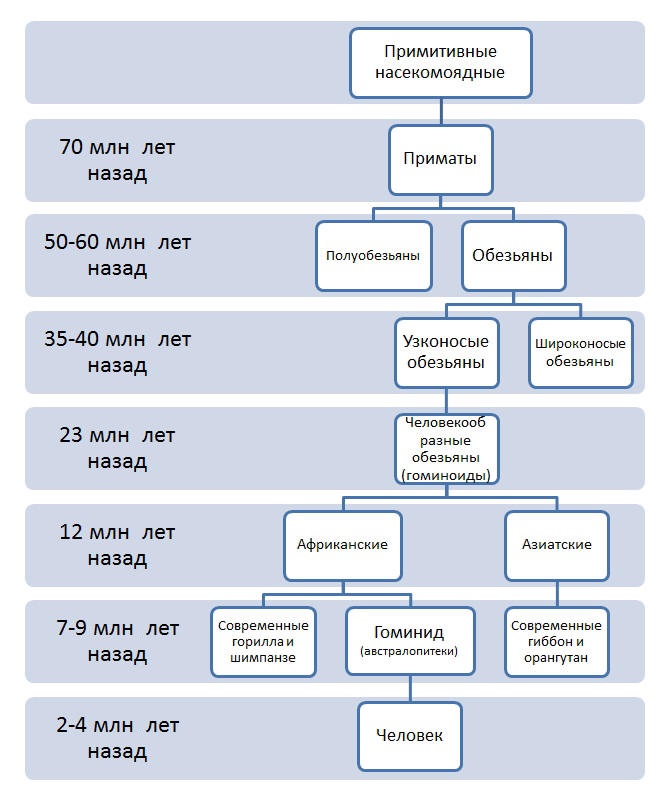
 |
|
Рис. 49. Эволюция человека |
Первые гоминиды были представлены разнообразными австралопитеками (рис. 49, а). Австралопитеки - многочисленная группа гоминид с объемом мозга менее 750 см3, они изготавливали каменные орудия труда, возможно, даже пользовались огнем.
Род человек отделяется от одной из ветвей австралопитеков в Юго-Восточной Африке примерно 2-4 млн лет назад (начало плейстоцена). К наиболее ранним находкам относится олдувайский человек, или человек умелый (Homo habilis) (рис. 49, б), который мало отличался от австралопитеков, но создал древнейшую орудийную культуру - олдувайскую. Ее основой была оббитая заостренная галька - чоппер. Объем головного мозга хабилисов достигал 650 см3. Хабилисы вытеснили с эволюционной сцены более примитивных австралопитеков, причем не гнушаясь и поеданием своих биологических родственников (разбитые черепа австралопитеков найдены на стоянках хабилисов). С возникновением гоминид начинается процесс социогенеза. Первой его фазой было первобытное стадо или первобытная орда.
Примерно 1,6 млн лет назад объем мозга у некоторых форм людей (человек прямоходящий, или Homo erectus) (рис. 49, в) превысил 750 см3 - этого оказалось достаточно для появления речи и формирования зачатков общества. Начинается расселение человека из Африки и проникновение в высокие широты (питекантропы, синантропы, гейдельбергский человек). Практически все эти ветви оказались тупиковыми. Homo erectus сумел достичь многого: он научился пользоваться огнем, создал ашельскую культуру (культуру ручных рубил). Несмотря на наступивший ледниковый период, он расселился на огромной территории, впервые покинув Африканский континент и постепенно покорив весь Старый Свет.
Примерно 400-500 тыс. лет назад формируется вид человек разумный (Homo sapiens) (рис. 49, г) с объемом мозга до 1500 см3. Один из подвидов - неандерталец (Homo sapiens neandertalensis) - создал богатейшую мустьерскую культуру каменных орудий.
В эпоху максимальных оледенений (60-200 тыс. лет назад) формируется непосредственный предок современного человека - кроманьонец (Homo sapiens sapiens fossilis) (рис. 49, д). Главным преимуществом кроманьонца перед неандертальцем был переход от первобытного стада к семье и родоплеменной организации. Примерно 40-30 тыс. лет назад неандертальцы вымирают, а человек современного типа (Homo sapiens sapiens) (рис. 49, е) формирует расы и заселяет все материки Земли.
Человек является высшим, но не обязательно последним звеном эволюции жизни на Земле. По мнению ряда ученых, человек как биологический вид, вследствие углубляющихся противоречий между состоянием окружающей среды с возможностями и потребностями человека находится в кризисе, который может завершиться постепенным его вырождением. В этих условиях, по их мнению, началось формирование нового биологического вида, который, в конечном счете, вытеснит человека и займет его место на планете. Другие исследователи полагают, что биологическая эволюция человека полностью прекратилась, сменившись эволюцией социальной. Но так или иначе эволюция всегда шла таким образом, что в ее процессе возникали все более сложные системы. Наиболее сложной из них является мозг человека. Именно мозг генерирует все рациональные схемы, к которым человек приспосабливает себя, свою деятельность и окружающий его мир.
11.5. О современной дискуссии по поводу теории эволюции
В современных дискуссиях вокруг проблем теории эволюции считается почти общепризнанным, что теория эволюции сталкивается с серьезными трудностями в объяснении явлений живой природы и не в состоянии решать возникающие здесь проблемы. К числу таких проблем можно отнести, в частности, реальность видообразования и макроэволюции, возможность прогрессивного усовершенствования в эволюции, механизмы формирования и трансформации сложных структур в эволюции, целесообразность строения живых организмов. Стереотипные представления об этих разделах теории эволюции широко используются современными креационистами с целью дискредитации науки. Между тем обсуждение имеющихся данных позволяет утверждать, что в решении каждого из упомянутых вопросов теория эволюции дает вполне удовлетворительные объяснения наблюдаемым фактам.
В настоящее время реальность эволюции уже не подвергает сомнению никто из серьезных ученых, хотя существует несколько конкурирующих теорий, каждая из которых предлагает свой вариант развития событий. В этом отношении эволюция аналогична гравитации. Существует несколько теорий гравитации - закон всемирного тяготения Ньютона, общая теория относительности, и, в один прекрасный день, возможно, появится универсальная теория. Однако существует факт тяготения - если вы уроните любой предмет, он упадет. Подобно этому существует факт эволюции, несмотря на то, что споры ученых по частным вопросам теории продолжаются.
Английский биолог Р. Докинз писал: «В наши дни теория эволюции вызывает примерно столько же сомнений, сколько теория о вращении Земли вокруг Солнца». Однако до сих пор число ниспровергателей этой теории не уменьшается. В частности, утверждается, что очевидная целесообразность строения и поведения живых организмов не могла возникнуть вследствие естественного отбора. В книге «Слепой часовщик» Докинз пишет: «...Мы знаем о животных и растениях гораздо больше, чем Дарвин, но и по сей день мне не известен ни один пример, когда сложный орган не мог бы появиться в результате целого ряда последовательных и незначительных модификаций. Я не думаю, что такой пример когда-либо будет обнаружен. Если же это произойдет - и такой орган действительно будет органом высокой степени сложности, а ...понятие «незначительности» будет определено в точных научных терминах - то я перестану верить в дарвинизм».
Однако ряд современных ученых утверждает, что на молекулярном уровне уже открыто немало таких неупростимых сложных систем и что по мере развития науки их открывают все больше и больше.
В конце ХХ века книга американского профессора Майкла Дж. Бихи под названием «Чёрный Ящик Дарвина» с подзаголовком «Биохимический вызов эволюционной теории» является предметом оживлённой дискуссии, нередко в весьма эмоциональной форме, и нередко выходящей далеко за рамки её казалось бы сравнительно специальной темы. Рассматривая данные современной биохимической науки, Бихи утверждает, что они несовместимы как с эволюционной теорией Дарвина, так и с её более современными модификациями, часто именуемыми нео-дарвинизмом.
Если обсуждать историю жизни на Земле, то следует рассмотреть две стадии, на каждой из которых события были обусловлены двумя разными принципами. На первой стадии процессы химической эволюции на древнейшей Земле привели к образованию первой живой клетки из неорганических материалов. На второй стадии потомки этой живой клетки развивались в разных направлениях, порождая многообразие жизни на планете, которое мы наблюдаем сегодня. На этой стадии развитие определял принцип естественного отбора.
О теории эволюции можно сказать, что биологические организмы созданы «слепым часовщиком», т. е. бессознательными дарвиновскими процессами мутации и отбора. В этом случае имеется своеобразная лестница со множеством ступенек, ведущая от первого организма до самого сложного: должна существовать непрерывная серия жизнеспособных промежуточных форм. Каждая последующая ступенька, ведущая к большей сложности, должна означать большую приспособляемость (в том, что касается потомства), в то же время как разрыв между ступеньками должен быть достаточно невелик, чтобы его можно было преодолеть посредством случайных мутаций.
Бихи рассматривает вопрос о формировании зрительных функций глаза: «Когда свет касается сетчатки, фотон вступает во взаимодействие с ретинальной молекулой H-cis, которая в течение одной пикосекунды трансформируется в трансретинальную молекулу. Изменение в форме ретинальной молекулы влечет за собой изменение в форме белка родопсина, с которым эта молекула прочно связана. Метаморфоз протеина изменяет его поведение. Теперь он уже называется метародопсин II и прикрепляется к другому типу белка - трансдуцину. До столкновения с метародопсином II у трансдуцина была маленькая молекула (GDP). При взаимодействии трансдуцина и метародопсина II эта молекула отпадает, а родственная ей, но принципиально отличная молекула (GTP) прикрепляется к родопсину».
Бихи называет такие (и аналогичные) системы системами неупростимой сложности. Такую систему невозможно выстроить путем последовательного добавления элементов, т. к. полезная функция каждого элемента возникает только тогда, когда он интегрирован в законченную систему. Но в этом случае организмы (и Вселенная) являются продуктом рационального сознания, т. е. построены по замыслу (Бога?). Ключевой аргумент Бихи состоит в том, что ни в каких научных журналах не встретишь подробных, подверженных проверке сценариев того, как невероятно сложные биохимические системы могли бы сформироваться под воздействием дарвиновских процессов.
Однако концепция «неупростимой сложности» пока остается лишь концепцией, поскольку, сколько бы неупростимой не казалась сложная система, думается, что такая неупростимость представляется лишь на современном этапе развития науки. Но это лишь отражает сложность изучаемых систем: каждое ядро содержит закодированную базу данных, превосходящую по информационному содержанию Большую Советскую Энциклопедию. И эта информация относится к одной клетке, а не к состоящему из клеток организму. Сложность анализа биологических объектов состоит еще и в том, что по конечному результату (строению современной клетки) мы должны восстановить путь и процесс создания (эволюции) живой клетки. На это и направлены усилия ученых.


